
The painting, by G. Paselk, is an interpretation of the Precambrian mural at the Natural History Museum of the Smithsonian Institute. The pool in the foreground represents a hydrothermal vent with mineral deposits around its rim. Similar vents may have incubated the origin of life on Earth. The yellow mounds in the larger water body are stromatolites.
4,570 to 542.0 Million years ago
Richard Paselk*
Plate Tectonic Reconstruction
Over the four billion years of the Precambrian the Earth's surface features changed vastly. Beginning as glowing ball of molten rock, Earth cooled to the pre-plate tectonic realm of the Archean. This was followed by two billion years of plate tectonics during the Proterozoic.
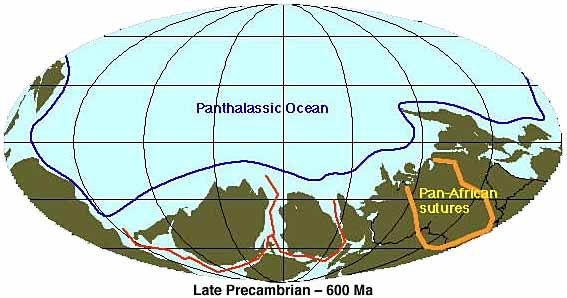
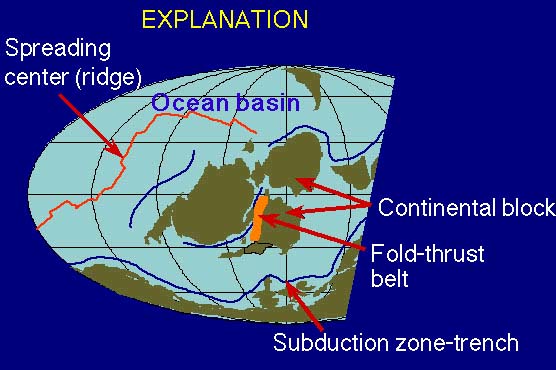
These maps of major tectonic elements (plates, oceans, ridges, subduction zones, mountain belts) are used with permission from Dr. Ron Blakey at Northern Arizona University. The positions of mid-ocean ridges before 200 Ma are speculative. Explanation of map symbols.
{kind=link}
The Precambrian encompasses nearly 90% of the history of the Earth and around a third of the history of the Universe. The Precambrian begins with the formation of the Solar System 4.57 billion years ago (bya) and extends to the beginning of the Cambrian 540 million years ago (Mya, or 0.54 bya). Over this immense time span the Solar System condensed from a cloud of dust and gas, the Earth was violently accreted from the collisions of smaller bodies, organic molecules were formed, and life originated from the resulting soup. In a very short period (geologically speaking) the progenitor cell of all modern life evolved and then diverged into the three great domains of modern life on Earth: the bacteria, the archea (bacteria-like organisms often living in extreme environments), and the eukarya (protists, fungi, plants and animals). Over the next three billion years these three domains of life diversified into the incredible variety of organisms appearing in the "Cambrian explosion."
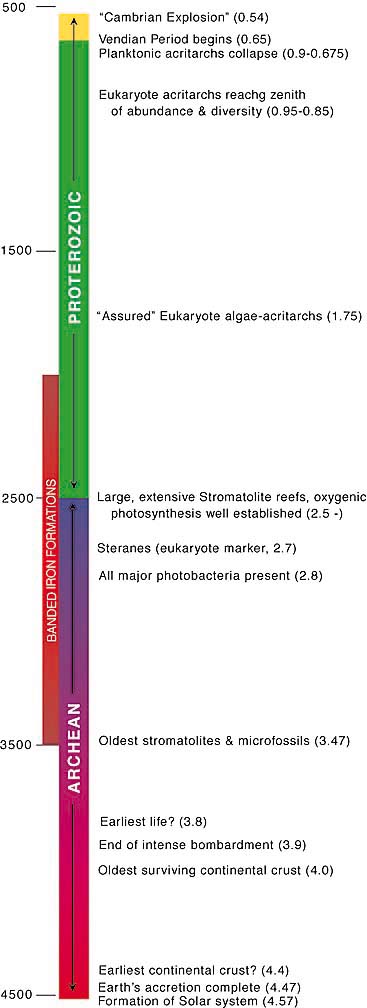
Essentially all of the great inventions of life were made during the Precambrian, including the ability to capture the energy of sunlight, the creation of an oxygen atmosphere, metabolism, the vast repertoire of biosynthetic products, sexual reproduction, the ability to detect light, communication by chemical signals, the metazoan body plans, and vascular plants all appear during this time. In fact the marvelous diversity of living organisms seen in the past five hundred million years is essentially just variations on themes already invented - refinements if you will of the inventions of the Precambrian. For those who wish to learn more about the Precambrian a bibliography is available via this link.
Meteorites
Most meteorites were formed within about 100 million years of the formation of the solar system. They are the oldest objects on Earth and they tell us about the conditions in the solar system when the Earth was just being condensed out of the solar nebula. Most meteorites are fragments of asteroids resulting from collisions in the asteroid belt. The most primitive meteorites are undifferentiated, that is they have never been melted and their component materials allowed to separate under gravitational attraction or recombine chemically. Such meteorites may have formed on the surface of an asteroid or been part of an asteroid too small to melt during its accretion. Meteorites originating from the interiors of large asteroids, on the other hand show varying degrees of differentiation, depending on the size of the asteroid, and thus how long it may have remained in a molten state.

- Chondrite (Stony Meteorite) with fusion crust (4.56 billion years old). Chondrites are characterized by chondrules - small spherules of melted rock which condensed out of the early solar nebula and then accreted into asteroids. This specimen is made up of Olivine ([Mg,Fe]2SiO4)-hypersthene([Mg,Fe]SiO3] and represents the primitive material of solar system with little post-accretional differentiation. (Fell in Holbrook, Arizona in 1912.)
- Stony Iron Meteorite (Pallasite polished section; about 4.5 billion years old): Pallasites have a continuous iron-nickel (Fe-Ni) alloy matrix with centimeter sized olivine crystal inclusions. These meteorites appear to come from a partially melted and differentiated asteroid (or part of an asteroid), and are thus slightly younger than the chondrites. (Found in Brenham, Kansas, 1882.)
- Iron Meteorite (octahedrite, 7% Ni Fe-Ni alloy; about 4.5 billion years old): These meteorites appear to come from a differentiated asteroid (or part of an asteroid), and are thus slightly younger than the chondrites. (Canyon Diablo Meteor crater, Arizona.) Specimen on Please Touch shelf.
{kind=link}
{kind=link}
{kind=link}
Precambrian Rocks and Minerals
One of the great stories of the Precambrian is the evolution of an oxygen atmosphere. Oxygen in the atmosphere not only resulted in one of the greatest ecological crises in Earth's history, it also was a necessary precursor to the evolution of multicellular animals (metazoans). Of course an oxygen atmosphere is a direct consequence of living organisms - it could not exist with just geologic/atmospheric origins. A direct result of the production of oxygen is the conversion of iron pyrite (FeS2) and oxygen (O2) into hematite (Fe2O3) and gypsum (CaSO4.2H2O).
So when did life first begin producing oxygen in significant amounts? The earliest records of cells are microfossils of cyanobacteria from about 3.5 bya, with recognizable stromatolite fossils found in Australia by 3.45 bya. Since all modern cyanobacteria produce oxygen by photosynthesis it is widely thought that oxygen production must date back to at least this time. Mysteriously, the geological evidence (banded iron formation, sulfide vs. sulfate minerals, cerium vs. cerium oxide deposition in paleosols etc.) indicate that oxygen didn't rise above trace levels until over a billion years later (2.3 bya). A variety of explanations have been offered, for example, the oxygen may have been consumed by early geological processes. But like much in the Precambrian, the true cause remains unknown.
A variety of specimens in our collection document the change to an oxygen atmosphere.

- Iron pyrite can only be formed in an oxygen-free environment, and is broken down to iron oxides and sulfate when oxygen is present during weathering and transport by streams and rivers.
- Iron Pyrite crystals such as this specimen occur in metamorphic and igneous source rocks.
- Iron pyrite and uraninite** are two minerals that decompose so rapidly in the presence of free oxygen that they are rarely if ever deposited in recent sediments. Both occur commonly in Archean detrital sediments such as the:
- Witwatersrand conglomerate (Polished slab, about 2.7 billion years old) in our collection. Note the rounded pebbles showing a stream-detrital deposit, while the presence of the pyrite flakes and uraninite indicate deposition under anaerobic atmospheric conditions. These ancient placers are mined for gold in South Africa to depths of 4 km. They have produced almost half the world's gold and some uranium.
- Pyrite Pebbles (2.3 billion years old, Transvaal, South Africa). Note the pyrite crystals have been rounded by tumbling in an oxygen-free environment.
- Iron Pyrite Crystals in Oil Shale.: Massive amounts of oil shale were deposited during the Precambrian. Comparisons of the carbon/sulfur isotopes in this shale to modern marine deposits indicate that this carbon is of biological origin. Iron pyrite crystals are often found in Archean and Proterozoic Precambrian shales, indicating the seas of this time were rich in soluble ferrous ion (Fe2+) and a source of sulfide ion. The only source of sulfide ion in shale would be from the metabolism of sulfate to sulfide by sulfate-reducing bacteria. Such bacteria must therefore have evolved and been active by at least 2.7 bya, when such deposits become common.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
- Banded Iron-Formation (BIF) constitutes the majority of the world's iron deposits. Most commonly these deposits consist of alternating layers of black hematite and chert. In our specimens the chert is colored red with iron oxide impurity coated granules.
BIF deposits are a direct result of oxygen release by Precambrian microbes. During much of the Precambrian the Earth's surface waters and atmosphere were anoxic (oxygen-free) so that iron would exist mostly in its reduced (ferrous or Fe2+) form. Vast quantities of ferrous iron entered the ocean surface through volcanic action, upwelling, and run-off. Photosynthesis by cyanobacteria in the surface waters produced oxygen which reacted with ferrous iron to give the much less soluble ferric iron (Fe3+), precipitating out iron hydroxide (rust). Seasonal and/or biological cycles resulted in intervening periods when iron or oxygen were not as available resulting in the interlayered chert (microcrystalline quartz precipitate).
- Banded Iron-Formation chunk.
- Banded Iron-Formation Slab, about 2.5 billion years old, Australia. Specimen on Please Touch shelf.
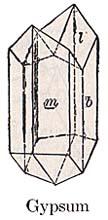
- Cleavage fragment of gypsum. Gypsum is soluble in water so is largely recycled into younger evaporite sediments. Some survives, especially as anhydrite CaSO4, in Precambrian rocks where its presence demonstrates the presence of oxygen due to photosynthesis.
{kind=link}
{kind=link}
**Uraninite is rapidly oxidized to higher uranium oxides. Though more complex in reality, the reaction can be symbolized as:
2 UO2+ O2-> 2 UO3
Precambrian Microbial Life
The earliest evidence of life comes from chemical fossils formed only 100 million years after the end of the Hadean period of intense bombardment by meteorites and planetesimals (3.8 bya), during which huge impacts by planetesimals would periodically vaporize the oceans and sterilize the Earth. This earliest life is inferred because the carbon 13:carbon 12 ratio (13C/12C) of carbonaceous deposits is characteristic of biochemical processes which preferentially use 12C. The earliest records of cells are microfossils of cyanobacteria from about 3.5 bya, with recognizable stromatolite fossils found in Australia by 3.45 bya.

- Shallow sea bottoms appear to have been covered by mats of cyanobacteria (engraving) due to a general lack of grazing animals prior to the Cambrian. A cast of a bacterial mat (sometimes called "Elephant skin") from the White Sea region of Russia is displayed.
- Stromatolites are the dominant fossil type for most of the Precambrian, with the oldest identified examples going back at least 3450 million years. During the Precambrian stromatolites formed reefs comparable in extent and magnitude to the great coral reefs of recent and modern times. Stromatolites became rare during the Cambrian with the advent of multicellular grazing animals from which they have little natural protection. Modern examples occur only in protected environments such as high salt lagoons where animals don't exist (e.g. the famous Shark's Bay examples in Australia), and in sand fields (such as in the photograph to the left taken in the Bahamas) where they are periodically covered in sand, denying access to grazing animals. Stromatolites are columnar or dome shaped finely layered structures made up of mineral deposits resulting from bacterial community growth. In modern stromatolites the living portion of the stromatolite is a complex ecological community dominated by filamentous cyanobacteria on the thin, surface, photosynthesizing, layer, with thicker, anaerobic layers underneath. As the bacteria grow sand and other particulate matter is trapped in the mat, along with calcium carbonate deposits, to make up the mineralized structure. The bacteria migrate upward to maintain access to light and food, gradually building the mineralized stromatolite below. Ancient stromatolites have fossil remains of cyanobacteria-like organisms which appear identical to their modern descendants.
{kind=link}

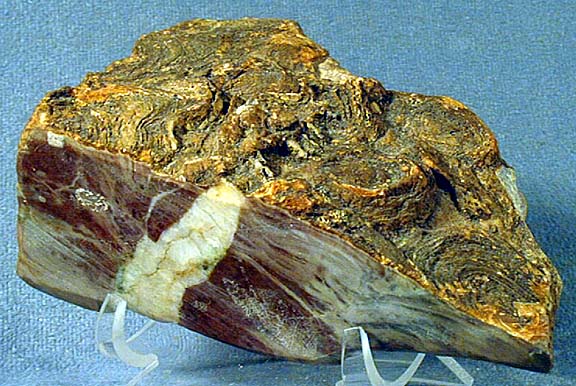
- Stromatolite (Proterozoic, 1200 Mya, Mount Nelson Formation, British Columbia)
- Stromatolite (Proterozoic, ca. 1800 Mya, Biwabik Formation, Minnesota). The typical layered structure of stromatolites is obvious in this specimen. The structure of this specimen shows something of the growth and ecology of these biostructures. Photosynthetic cyanobacteria on the surface released oxygen that then precipitated red iron oxide during the day, adding layer after layer (see the closeup image showing the fine structure of these layers). As the layers build up the buried layers the oxygen was consumed and anerobic bacteria thrive, some metabolizing iron oxide and sulfur to create crystals of iron pyrite seen as golden flecks deeper in the stromatolite. Two crystals are also visible in the closeup. The white quartz layer is a later intrusion as the rock was buried and fractured over time.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Multicellular Animals - the Ediacaran Fauna
Multicellular animals first become significant life-forms, leaving the earliest clear fossil evidence, at the beginning of the Vendian, about 0.6 bya (600 Mya). The Ediacaran fauna of the Vendian Period is represented by the casts in our collection from the famous fossil beds of the Flinders Ranges of South Australia, the White Sea beds of Northern Russia, and the beds in Charnwood Forest of Leicestershire, England. The Ediacaran fauna probably represent the first ecosystem in which multicellular organisms play a significant role.
The Ediacaran fauna represent a mystery however - with the exception of coelenterates (medusa and anemones), these organisms are not clear ancestors of those which followed them in the Cambrian. In addition, modern DNA-based sequence information predicts that today's phyla split much earlier than the Vendian, perhaps as much as a billion years ago. So where did the organisms of the Cambrian, and our modern phyla, come from?
As with many later Periods the Vendian probably ended with a mass extinction, in this case associated with an ice age. The extent of the Vendian extinction event may have even exceeded later events, such as the Permo-Triassic crisis, when 90% of marine species vanished. In this case the loss of the Ediacaran fauna allowed the rapid diversification of bilaterian animals to fill the empty niches. Recent work has shown that the Cambrian Explosion was not only a phenomena of bilaterian animals, it was an ecosystem-wide diversification of organisms including bacteria, protists, algae, and sponges as well.
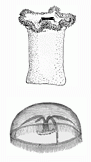
Radiata: The Radiata (coelenterata) include today's Cnidaria (hydras, jellies, sea anemones, and corals) and Ctenophora (comb jellies) and are characterized by radial symmetry. They probably represent the earliest multicellular animals with true tissues, and strongly dominate the Vendian fauna around the world. Fossils of Radiata, impressions since they have no hard tissues, are common in the Vendian Period.
The Radiata are represented by two body-plans: polyps (e.g. anemones and corals) and medusa (jellies).
- Polyps: The burrowing sea anemone (Niemiana), is thought to represent the earliest and most primitive Radiata. Note that they have no tentacles, as seen the the group in our cast from the White Sea deposits of Russia, and unlike modern examples as in the engraving. These organisms also had an undifferentiated body cavity and reproduced asexually.
- Medusa: Medusa-like fossils have been found as old as 1200 Mya in Australia. As with the polyps they appear to be much simpler than modern medusa, such as the jelly in the engraving. The museum has a number of representative fossil casts of White Sea specimens courtesy of M. A. Fedonkin (Paleontological Institute, Moscow) and the University of California Museum of Paleontology:
- Cyclomedusa represents one of the earliest forms of medusa. It was a sedentary form, which may have evolved by a flattening of a polyp resulting in a concentric body plan. This simple organism reproduced asexually by fission. (White Sea deposits of Russia)
- Protodipleurosoma rugulosum another sedentary form of medusa, it appears to have reproduced asexually by budding. (White Sea deposits of Russia)
- Eoportita appears to be a more advanced form of medusa, incorporating both concentric and radial symmetry in its body plan. In the cast the radially arranged tentacles can be seen to be concentrically arranged in series around the oral cavity. As with Cyclomedusa, reproduction was asexual. (White Sea deposits of Russia)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
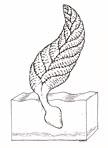
Colonial Forms: Some authors also consider a variety of other pinnate-shaped, fan-shaped, and comb-shaped organisms to be colonial representatives of the Radiata. Others consider them to be failed early experiments in animal biology, with no modern descendants. These organisms have no apparent mouths or anuses, thus they must have absorbed food, or harbored photosynthetic symbionts to provide them with energy. Examples of both modes of feeding exist in modern organisms. A variety of corals have never been observed to feed, rather existing on the photosynthetic products of symbiotic cyanobacteria. And the giant worms found around hydrothermal vents in the ocean have no mouths or anuses. Rather they harbor symbiotic bacteria which use the methane, hydrogen sulfide and other reduced substances to generate energy for themselves and their hosts. The original ink drawing by Rachel Rogge at the left is an artists representation of a living Charnia masoni commissioned by the museum (©2002 Cal Poly Humboldt NHM).
Casts of Charnia masoni fossils from two localities are on display:
- The White Sea region, Russia (positive and negative impressions), courtesy of M. A. Fedonkin (Paleontological Institute, Moscow) and the University of California Museum of Paleontology.
- Charnwood forest, Leicestershire, England, Two specimens, second on Please Touch shelf
{kind=link}
{kind=link}

Trilateria: A variety of Vendian organisms exhibit trilateral radial symmetry. The relationship of these animals to modern organisms is controversial. Many paleontologists feel they represent early experiments in animal biology which failed, becoming completely extinct and leaving no modern representatives. The original ink drawing by Rachel Rogge at the left is an artists representation of a living Tribrachidium commissioned by the museum (©2002 Cal Poly Humboldt NHM).
- The cast of Tribrachidium heraldicum in our collection is from the famous Ediacara fossil beds of Australia courtesy of James Gehling. It may represent a more advanced form of medusa, with more similarity to modern cnidarians than with other Vendian medusa.
- Pteridinium is a frond-like organism with three-fold symmetry, which may indicate a relationship to the Tribrachidium discussed above. On the other hand its frond-like body plan could argue for a closer relationship to Charnia, and a colonial nature. The Museum's cast of a White Sea specimen is courtesy of M. A. Fedonkin (Paleontological Institute, Moscow) and the University of California Museum of Paleontology.
{kind=link}
{kind=link}

Bilateria: The bilateria (animals with bilateral symmetry such as the majority of living forms) are represented by two types of Precambrian fossils: trace fossils and impressions. Trace fossils of burrows and tracks are perhaps the best evidence for the existence of animals ancestral to modern forms in the Precambrian. Unlike the various fossils represented in the Vendian impressions, these animals would have possessed the key characteristic of most animals - mobility. Unfortunately there are no known fossils of these organisms themselves, so we can't know who made these tracks and burrows.
Among the best known and common impressions of animals thought to represent bilateria are the fossils of Dickinsonia. These discoidal, segmented animals are thought by some experts to represent early annelid worms, while others feel they are unique Ediacaran species with no descendants. The Dickinsonia varied greatly in size, from small coin-sized to doormat sized. Some impressions are surrounded by smoothed areas indicating the organisms expanded and contracted while maintaining their shapes and segmentation numbers. Some specimens appear to have orifices which have been interpreted as a mouth and an anus at the ends of the central ridge, though others dispute this interpretation. Perhaps future specimens will enable us to determine the true relationships of these and other Ediacaran animals, but currently we just don't know. The original ink drawing by Rachel Rogge at the left is an artists representation of a living Dickinsonia commissioned by the museum (©2002 HSU NHM). The museum has two casts of Australian specimens provided courtesy of James Gehling:
{kind=link}
{kind=link}
The engravings are from Dana, James D. (1870) Manual of Geology, Dana, Edward S. and William E. Ford (1922) A Textbook of Mineralogy, and Strasburger, Eduard et. al. (1920) A Text-book of Botany.
*The Precambrian exhibit and web site were researched and created over a two year period ending in August of 2002. Though I, Richard Paselk will take blame for any errors etc., I wish to acknowledge the aid and generosity of a number of individuals. I would like to thank Jere Lipps (UCMP), Don Garlick (Humboldt emeritus), Ken Aalto (Humboldt) and John Longshore (Humboldt emeritus) for helpful discussions on the Precambrian and geology in general. Holly Harvey has been my Illustrator& Photoshop guru. Melissa Zielinski has encouraged me, edited both web pages and display legends, been invaluable in obtaining specimens, and just generally been a great friend and colleague in doing this project. Finally, my wife Gail has not only contributed her artwork and layout skills, she has also been a supportive and helpful companion throughout the difficulties and frustrations of this seemingly small but as it turned out long and challenging project.
Nearly all of the specimens on display were provided as gifts to the museum courtesy of the individuals listed below:
- Don Garlick, Humboldt Geology Dept. (emeritus) generously provided the three meteorites and the various mineral specimens.
- Ken Aalto, Humboldt Geology Dept. provided the stromatolite specimens and photo of Bermuda stromatolites.
- Jere H. Lipps, University of California Museum of Paleontology, arranged for the casts of Ediacaran life from specimens provided by M.A. Fedonkin, Palaeontological Institute, Russian Academy of Science (White Sea specimens), and James Gehling, University of South Australia (Australian specimens).
- Mihai Tomescu, Humboldt Biology Dept. generously provided the iron formation stromatolite.
Bibliography
Knowledge about the precambrian has changed rapidly in recent years. Two articles summarizing some recent work appeared in National Geographic in March and April of 1998. Written by science writer Richard Monastersky, both are written and magnificently illustrated in typical National Geographic style. The first focuses on the origin and early evolution of single-celled life, while the second focuses on the Ediacaran fauna.
- Monastersky, Richard (March 1998) "The Rise of Life on Earth." National Geographic 193 (3) pp 54-81.
- Monastersky, Richard (April 1998) "Life Grows Up." National Geographic 193 (4) pp 100-115.
For material in greater depth than provided in the National Geographic articles above I recommend the books below and the articles in Major Events in the History of Life, edited by J. William Schopf.
- Fedonkin, Mikhail A., et. al. The Rise of Animals: Evolution and Diversification of Animalia. The Johns Hopkins Press, Baltimore (2008).
- Knoll. Andrew H. Life on a Young Planet—the First Three Billion Years of Evolution on Earth. Princeton University Press, Princeton (2003).
- Schopf, J. William (1992) "The Oldest Fossils and What They Mean." in Major Events in the History of Life. (J. William Schopf, ed.) Jones and Bartlett Publishers, Boston.
- Runnegar, Bruce (1992) "Evolution of the Earliest Animals." in Major Events in the History of Life. (J. William Schopf, ed.) Jones and Bartlett Publishers, Boston.
The references below represent most of the articles I used in researching this display and the associated web pages. Most of the articles from Science and Nature are mini-reviews of the latest research written for a scientifically sophisticated, but non-specialist audience. All of the articles reference the specialist literatures.
Richard A. Paselk, Precambrian exhibit designer/author.
- Des Marais, David J. (8 September 2000) "When Did Photosynthesis Emerge on Earth?" Science 289 pp 1703-5.
- Halliday, Alex N. (11 June 2000) "In the Beginning" Nature 409. pp 144-5.
- Holland, Heinrich D. (2 January 1997) "Evidence for Life on Earth More Than 3850 Million Years Ago." Science. 275. pp 38-9.
- Jørgensen, Bo Barker. (19 July 2001) "Space for hydrogen." Nature 412. pp 2869
- Knoll, Andrew H. and Sean B. Carroll. (25 June 1999) "Early Animal Evolution: Emerging Views from Comparative Biology and Geology" Science284. pp 212937.
- Kramers, Jan. (27 July 2001) "The Smile of the Cheshire Cat" Science 293. pp 619-20.
- Lipps, Jere H. and Philip W. Signor. (1992) Origin and Early Evolution of the Metazoa. Plenum Press, New York.
- Martin, M. W., D. V. Grazhdankin, S. A. Bawring, D. A. D. Evans, M. A. Fedonkin, and J. L. Kirschvink. (5 May 2000) "Age of Neoproterozoic Bilaterian Body and Trace Fossils, White Sea Russia: Implications for Metazoan Evolution." Science 288. pp 841-5.
- Meyerowitz, Elliot M. (22 February 2002) "Plants Compared to Animals: The Broadest Comparative Study of Development." Science 295. pp1482-5.
- Nisbet, Euan. (8 June 2000) "The realms of Archean life" Nature 405. pp 6256.
- Sleep, Norman H. (15 March 2001) "Oxygenating the atmosphere." Science 310. pp 3179.
- Rasmussen, Birger, Stefan Bengtson, Ian R. Fletcher, and Neal J. McNaughton (10 May 2002) "Discoidal Impressions and Trace-Like Fossils More Than 1200 Million Years Old." Science 296. pp 1112-5.
- (22 December 2000) Science. 290. p 2241
- Walker, James C.G., Cornelis Klein, Manfred Schidowski, J. William Schoppf, David Stevenson, and Malcolm R. Walter (1983) "Environmental Evolution of the Archean-Early Proterozoic Earth" in Earth's Earliest Biosphere. (J. William Schopf, ed.) Princeton University Press, Princeton. pp 276-283 [detailed minerology etc. of banded iron-formations]
©2002, Cal Poly Humboldt NHM | Last modified 28 October 2012